Humanin Peptide Structure: Inside a 24-Residue Mitochondrial-Derived Peptide
Humanin is a rare peptide read out of the mitochondrion's own genome rather than the cell nucleus. This explainer walks through its 24-residue sequence, its three-turn alpha-helix, the potent S14G research analog, and how its compact structure maps onto the receptors and intracellular partners researchers study it against.
by Research Assistant·
Almost every peptide your cells build is read out of the nucleus, the main library of genetic information. Humanin is one of the rare exceptions. It's transcribed instead from the tiny, separate genome tucked inside the mitochondrion. That odd address is the reason humanin peptide structure — a compact chain of just 24 amino acids — is the natural place to start if you want to understand why this mitochondrial-derived peptide has drawn so much research attention. The compounds discussed here are for research use only and are not intended for human or animal consumption. What follows is educational: it describes what has been seen in laboratory studies, not outcomes in people.
We'll move from the literal sequence of residues, to where in the genome humanin is encoded, to the alpha-helix it folds into, and finally to how that small shape engages the receptors and proteins researchers study it against.
The Amino Acid Sequence: What Humanin Is Made Of
Start with the chain itself. The full-length form of humanin is 24 amino acids long, and in single-letter code its sequence reads MAPRGFSCLLLLTSEIDLPVKRRA (Wikipedia: Humanin). If you've ever worked through reading a peptide's residue sequence before, the logic is identical here — each letter is one amino acid, read from the N-terminus on the left to the C-terminus on the right.
A few features jump out even at a glance. The chain opens with a methionine (M), the standard start residue. Its middle is strikingly hydrophobic: a run of leucines around positions 9 through 12 — that LLLL stretch — gives the peptide a greasy, water-avoiding core. And it closes on a cluster of basic, positively charged residues, lysine and two arginines (K-R-R-A), at the C-terminal tail. That pairing of a water-shy middle with a charged tail turns out to shape how the peptide behaves, a theme we return to once we look at its folded form (Gong et al., 2014, PMC3705736).
Where Humanin Comes From: A Mitochondrial-Derived Peptide
So what is a mitochondrial-derived peptide? In short, it's a peptide whose instructions live in the mitochondrion's own DNA rather than in the cell nucleus. Mitochondria are best known as the cell's power plants, and their compact genome was long assumed to encode only a fixed set of housekeeping molecules.
The MT-RNR2 / 16S rRNA origin
Humanin is encoded by a short open reading frame — roughly 75 base pairs — nested inside the MT-RNR2 gene, which mainly produces the mitochondrial 16S ribosomal RNA (evolutionary analysis, PMC10465549). Put another way, a small protein-coding message hides inside a gene scientists had filed away as purely structural RNA. When it was described, humanin stood out as the first new peptide identified in the mitochondrial genome since that genome was fully sequenced back in 1981 (PMC3705736).
How it was discovered
The peptide was first reported in 2001 by the Nishimoto laboratory. The team was screening surviving neurons from an Alzheimer's-disease brain, hunting for factors that let some cells resist amyloid-beta-associated cell death — and the gene they pulled out encoded humanin (PMC3705736; Kim et al., 2022 review). That origin story is a piece of research history, not a claim about treating any condition. But it does explain why humanin's structure got such close scrutiny from the very start.
Two Isoforms: Why the Translation Site Changes the Length
One quirk of humanin is that it has no single fixed length. Because the peptide's message can be read by two different pieces of cellular machinery, it shows up in two closely related forms (PMC3705736).
A 24-amino-acid form appears when the message is translated in the cytoplasm, using the standard genetic code — this is the MAPRGFSCLLLLTSEIDLPVKRRA sequence above.
A shorter form of about 20 to 21 amino acids appears when the message is translated inside the mitochondrion, which runs a slightly different genetic code and trims residues from the C-terminal end (Wikipedia: Humanin).
Both versions have been reported as biologically active in laboratory studies. Humanin also lacks a conventional secretion signal, yet it can act as its own signal peptide — part of how researchers explain its appearance both inside and outside cells (PMC3705736). A review of the molecular structure adds that some species carry even longer variants, a point we pick up under conservation (Kim et al., 2022).
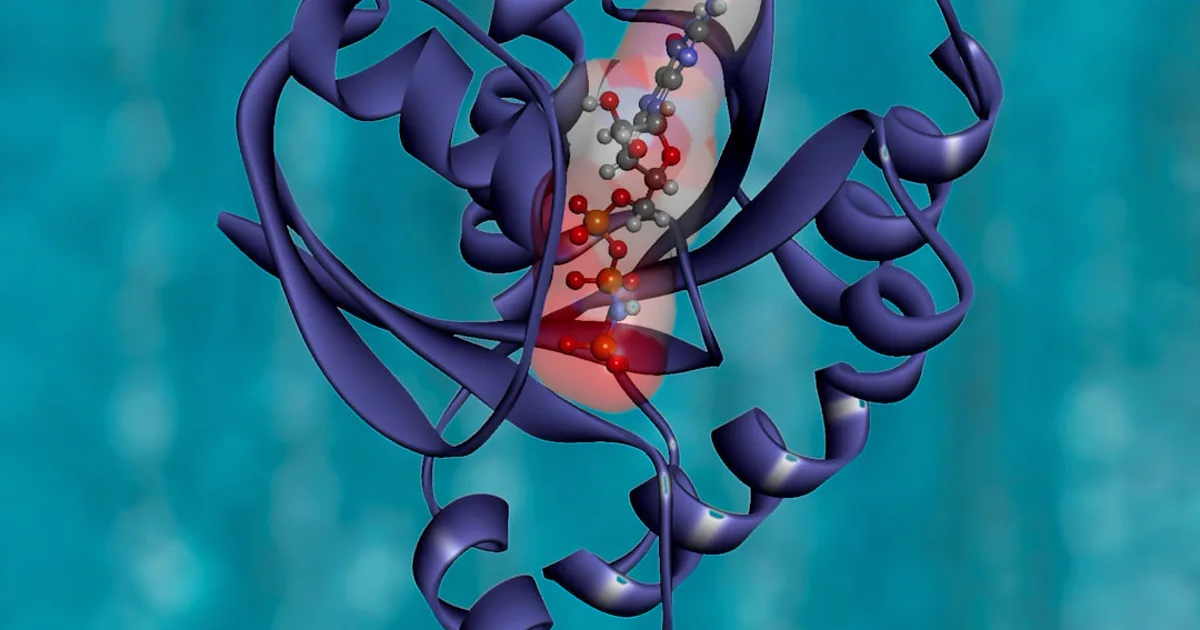
Secondary Structure — The Three-Turn Alpha-Helix
Primary sequence tells you the order of residues. Secondary structure tells you the shape they settle into. In solution, the humanin chain folds into a short three-turn alpha-helix with no internal symmetry (Wikipedia: Humanin). An alpha-helix is the familiar corkscrew coil of protein chemistry, and if you want the wider picture of how a peptide's shape relates to its function, humanin makes a tidy case study.
What makes the shape useful is that specific residues line up along the helix. Mutational studies — where researchers change one residue at a time and watch what breaks — flag a handful of positions as important for humanin's activity in cell-culture models: proline 3, serine 7, cysteine 8, leucine 9, leucine 12, threonine 13, and serine 14 (PMC3705736). Cysteine 8 is especially versatile in these studies, feeding into more than one of the peptide's laboratory interactions. Read together, those residues cluster into the functional face of the little helix.
The S14G Analog and Structure–Activity Relationships
If serine 14 is one of the load-bearing residues, what happens when you change it? That question is the origin of humanin's most famous laboratory variant. Swap the serine at position 14 for a glycine and you get an analog written as S14G, or HNG, often called "Humanin-G" (Yen et al., 2013, PMC3641182).
That single-residue change has an outsized effect. Across several assays, the S14G analog has been reported as up to roughly 1000 times more potent than the natural peptide in cell-protection experiments (Wikipedia: Humanin). It's a clean illustration of a structure–activity relationship — the idea that swapping one amino acid can sharply shift how a peptide behaves — and it's why S14G, not the wild-type sequence, is so widely used as a research tool. The same principle drives how researchers study other short signaling peptides, where small edits to a compact chain produce large changes in activity.
How Structure Maps to Receptor Binding
A compact, well-defined helix is a molecule that can dock onto specific partners, and humanin's structure supports interactions on both sides of the cell membrane (PMC3641182).
Outside the cell, humanin acts as a ligand for two receptor systems: the seven-transmembrane formyl peptide receptors (FPRL1 and FPRL2) and a three-part, cytokine-like receptor complex built from CNTFR, WSX-1, and gp130. A formyl group tacked onto the peptide's N-terminus — a modification typical of mitochondrially encoded proteins — has been reported to sharpen activation of these receptors (PMC3705736). Because it can engage more than one receptor system, humanin often comes up alongside peptides that engage more than one receptor.
Inside the cell, the same small peptide keeps different company. In laboratory studies it binds several pro-apoptotic proteins of the Bcl-2 family — Bax, tBID, and BimEL among them — and it also binds the protein IGFBP-3 at a region called its heparin-binding domain (PMC3641182). This dual reach, intracellular and extracellular, is a big part of why researchers describe humanin as a retrograde signaling molecule sent out from the mitochondrion.
Evolutionary Conservation and Species Variants
One way researchers judge whether a sequence matters is to ask how well evolution has preserved it. Humanin scores high. Analyses find strong synonymous codon bias and sequence conservation across vertebrates — a statistically robust signal of purifying selection (with a reported p-value of about 0.00063) that keeps the functional residues intact (PMC10465549).
Conservation doesn't mean the sequence is identical everywhere. Length varies by species: human humanin is 24 residues, while the rat version — sometimes called "rattin" — runs to 38, and some marsupials and hoofed mammals carry extended forms of 30 to 40 amino acids (PMC10465549). On top of the mitochondrial gene, the human nuclear genome carries 13 humanin-like homologs, named MTRNR2L1 through MTRNR2L13, several of which are transcribed in human tissues (Kim et al., 2022).
Why the Structure Draws Research Interest
Structure only matters because of what it lets a molecule do, and humanin has been characterized across a range of cell-culture systems. Take one well-documented example. Cultured retinal pigment epithelium (RPE) cells were placed under oxidative stress, and humanin added to the culture was associated with restored mitochondrial respiration, higher mitochondrial DNA copy number, and more mtTFA, a regulator of the mitochondrial genome (Sreekumar et al., 2016, PMC4811181).
In those same studies, humanin was linked to reduced Bax movement to the mitochondria and lower caspase-3 activity — the intracellular partners we met earlier — and to fewer senescence markers, in a way that depended on the STAT3 signaling pathway (PMC4811181). These are observations in cell culture, not outcomes in people. Still, they show why a compact helix able to touch both receptors and apoptosis machinery is such an interesting object of study.
Frequently Asked Questions
What is the amino acid sequence of humanin?
The full-length cytoplasmically translated form of humanin is 24 amino acids long, with the single-letter sequence MAPRGFSCLLLLTSEIDLPVKRRA. A shorter mitochondrially translated form of about 20 to 21 residues drops several residues from the C-terminus. Both forms have been reported as biologically active in research settings.
What does "mitochondrial-derived peptide" mean?
It means the peptide is encoded within the mitochondrion's own small genome rather than in the cell nucleus. Humanin comes from a short open reading frame nested inside the MT-RNR2 gene, which primarily encodes the 16S ribosomal RNA. This makes humanin one of the first recognized members of a class of small peptides read out from mitochondrial DNA.
What is the S14G humanin analog?
S14G is a synthetic version of humanin in which the serine at position 14 is replaced by glycine. Commonly written as HNG or "Humanin-G," this single-residue change has been reported to increase the peptide's cytoprotective potency by up to roughly 1000-fold in certain laboratory assays, which is why it is widely used as a research tool.
How is humanin's structure related to its function?
In solution the peptide folds into a short three-turn alpha-helix. Mutational studies point to a handful of residues — including positions 3, 7, 8, 9, 12, 13, and 14 — as important for its activity in cell-culture models, and its compact size lets it interact with both cell-surface receptors and intracellular protein partners. The research-use-only caveat applies throughout.
Putting It All Together
Humanin is a small molecule with an unusual biography: a 24-residue chain read out of the mitochondrion's own genome, folded into a three-turn alpha-helix, conserved across vertebrates, and able to touch both cell-surface receptors and intracellular proteins. The S14G analog shows how much a single residue can change, and the cell-culture literature shows why that compact structure is worth studying. If this was useful, the site's other structure explainers — on peptide shape, multi-receptor design, and related short peptides — are good next stops for seeing how sequence becomes function.
For research use only. Not for human or animal
consumption of any kind. The information in this article is for
educational purposes only and is not intended to diagnose, treat,
cure, or prevent any disease. The statements made have not been
evaluated by the U.S. Food and Drug Administration. These products
are NOT FDA APPROVED. Please consult with a licensed healthcare
professional before making any decisions regarding your health
or research.
Optides LLC is a chemical supplier. Optides LLC is not a
compounding pharmacy or chemical compounding facility as defined
under 503A of the Federal Food, Drug, and Cosmetic Act. Optides LLC
is not an outsourcing facility as defined under 503B of the Federal
Food, Drug, and Cosmetic Act.